Definitie en functie van meristemen
Lagere planten zoals varens en paardestaarten hebben als groeipunten
apicale cellen die de vorm van een omgekeerde piramide of schotel hebben.
Bij hogere planten vind je geen enkele apicale cel, maar groeizones die uit zeer veel cellen kunnen bestaan; ze worden
meristemen genoemd, waarin groepjes delende cellen voorkomen. Meristemen bestaan uit ongedifferentieerde cellen die het vermogen hebben om te delen en daarna te differentiëren (meristos betekent 'gedeeld' in het Grieks).
Type meristemen: topmeristemen, primaire meristemen en secundaire meristemen
Tijdens de vroege ontwikkeling van het plantembryo, dus nog voordat het zaad gekiemd is, worden aan de toppen van het worteltje en de spruit al twee
topmeristemen of
apicale meristemen aangelegd:
het
top-spruitmeristeem ook wel
apicale spruitmeristeem of
scheutmeristeem genoemd, leidt bovengronds tot de vorming van primaire meristemen waaruit primair weefsel in stengels en bladeren zich ontwikkelen,
het
top-wortelmeristeem leidt tot het ontstaan van de
primaire (wortel) meristemen waaruit primair wortelweefsel zich ontwikkelt.
Beide topmeristemen blijven gedurende de gehele ontwikkeling van de plant actief, zowel bij monocotylen (=eenzaadlobbigen) als bij dicotylen (=tweezaadlobbigen).
In wortel, stengel en bladeren worden de dezelfde drie typen primair meristeem en primair weefsel onderscheiden:
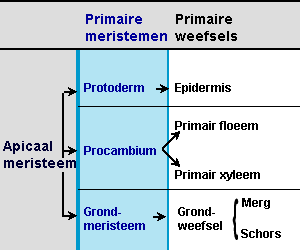
- Het protoderm (Pd) waaruit de epidermis ontstaat
- Het grondmeristeem (Gr) waaruit het grondweefsel ontstaat
- Het procambium (Pc) waaruit cellen ontstaan die zich sterk strekken en tot primaire vaatbundelcellen ontwikkelen: xyleemvaten (= houtvaten) voor het vervoer door de stengel van water van de wortel naar de bladeren toe en floeemvaten voor de verdeling in de plant van suikers die in de bladeren gemaakt zijn.
In de primaire meristematische zones vindt vooral
lengtegroei plaats. Dit gebeurt om te beginnen door gewone celdeling (
mitose). De nieuwgevormde cellen gaan vervolgens strekken, wat in aanzienlijke mate tot de uiteindelijk toename in lengte bijdraagt. Naast celstrekking vindt er ook uitzetting in de breedte oftewel expansie plaats.
Bij de meeste tweezaadlobbigen (dicotylen), maar niet bij eenzaadlobbigen (monocotylen), komt na de primaire of lengte-groei later ook nog secundaire groei voor. Deze vorm van groei, ook diktegroei of laterale groei genoemd (lateraal = naar de zijkant toe), vindt zijn oorsprong in secundaire (nieuw-gevormde) meristemen (meer over secundaire meristemen).
Voorbeelden: topmeristemen in embryo's en groeiende planten
| Topmeristemen in een plantembryo en een jong plantje |
 |
 |
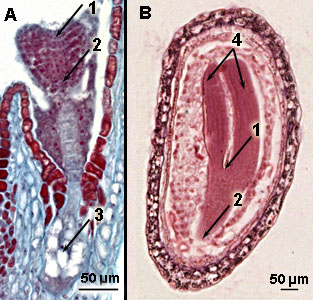
Meristemen in embryo's van het Herderstasje
A. Jong "hartvormig" embryo en B ouder "torpedo-vormig" embryo
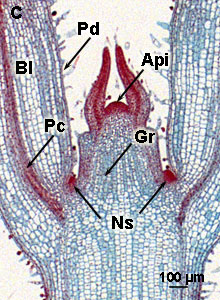
1 = top- of apicaal scheutmeristeem; 2 = top- of apicaal wortelmeristeem; 3 = hechtingscel of suspensor; 4 = zaadlobben of cotylen | Topmeristeem, oftewel apicaal scheutmeristeem, van de Japanse siernetel
Api = top of apicaal meristeem; Gr = grondmeristem; Pd = protoderm; Pc = procambium; Ns = Nieuwe zijscheuten;Bl = Blad
Aan de scheut ontstaan nieuwe zijscheuten (Ns) en bladeren (Bl) door celdeling (mitose) in de schors, vlak onder de epidermis, evenwijdig aan het oppervlak (periclinaal). Daarna vinden er ook delingen plaats dwars op het oppervlak (anticlinaal) en tenslotte ook in de epidermis (uitsluitend anticlinaal; zie hierbeneden een schets van anticlinale en periclinale delingen).
Naar een foto van het apicaal meristeem van de siernetel opgenomen met een elektronen microscoop |
| Organisatie van het apicale scheutmeristeem |
| Cellagen en -delingsrichtingen |
Canadese Waterpest (1,2,3)
(Eenzaadlobbige) |
Japanse Siernetel (4,5)
(Tweezaadlobbige) |
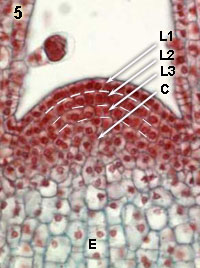
Apicaal scheutmeristeem bij de Canadese waterpest (eenzaadlobbige; linkse kolom) en bij de Japanse siernetel (tweezaadlobbige; rechtse kolom). In de laag cellen aan het oppervlak van het meristeem, vind je bijna alleen anticlinale delingen (delingsvlak loodrecht aan het weefseloppervlak; zie schets hieronder). Daardoor blijft de enkele aangesloten cellaag structuur gehandhaafd en wordt deze laag tunica of mantel genoemd. In de cellen onder de tunica, het zogenaamde corpus, vinden niet allleen anticlinale maar ook periclinale (delingsvlak parallel aan het oppervlak) en schuine delingen plaatst. Het volume van de stengel neemt daardoor sterk toe.
In de organisatie van tunica en corpus treden, afhankelijk van de soort, variaties op. Bij de waterpest, bestaat de tunica uit slechts één cellaag, maar bij de siernetel bestaat ze uit twee cellagen (aangegeven met L1 en L2). Met L3 is de zogenaamde initiale laag van het corpus (C) aangegeven. Pas na een aantal delingen gaan de nieuwgevormde cellen sterk expanderen (E), wat we dan zien als groei.
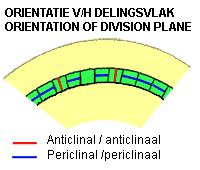
Anticlinale delingen: de nieuwe celwand ligt loodrecht op het weefseloppervlakte
Periclinale delingen: de nieuwe celwand ligt evenwijdig aan het celoppervlakte
| 
|

|

|
|
Groei en stengelbouw bij éénzaadlobbigen, tweezaadlobbigen en bij houtige Gymnospermen

Wanneer er meer zijstengels of zijtakken en dus ook bladeren komen is er ook meer vaat-, vul- en steunweefsel in de stengel nodig. Bij tweezaadlobbigen (voorbeeld: Japanse siernetel en Zonnebloem), maar ook bij houtige gymnospermen (voorbeeld: microscopische preparaten van een
dennenstam) ontstaat er een nieuwe delings- of cambiumlaag) die voor de dikte- of secundaire groei zorgt. De groei verloopt alleen in laterale richting (naar de zijkant toe); de stengel wordt dikker maar niet langer. (Meer over diktegroei in de stengel bij
secundaire meristemen).
Bij éénzaadlobbigen (voorbeeld: Canadese waterpest en Mais) is primaire groei de enige manier om het aantal cellen in de plant te vergroten. Palmbomen, die ook tot de eenzaadlobbigen behoren, kennen een vorm van schijn-diktegroei. Deze berust echter nog steeds op delingen in cellen afgeleid van het -primair- apicaal meristeem. Slechts een hele kleine groep éénzaadlobbige planten, waaronder
Dracaena, vertoont echte secundaire groei.
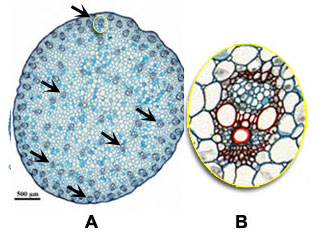
Kenmerkend voor éénzaadlobbigen is, behalve de afwezigheid van een vaatbundelcambium, dat de vaatbundels in de stengel verspreid liggen (zwarte pijlen in overzicht A bij mais), en dat elke vaatbundel een gesloten structuur vormt (aanwezigheid van een schede, zie B). Bij de tweezaadlobbigen zijn de vaatbundels gerangschikt in een niet aaneengesloten kring in jonge stengels (oranje pijlen in overzicht C bij de zonnebloem) en binnen de vaatbundel zijn xyleem en floeem gescheiden door een cambiumlaag (stippenlijn in D) waaruit verdere groei en differentiatie kan plaatsvinden.
| Bouw van de stengel: eenzaadlobbigen versus tweezaadlobbigen |
Eenzaadlobbige bijv. Mais (A en B) | Tweezaadlobbige, bijv. Zonnebloem (C en D) |
| In eenzaadlobbigen komen vaatbundels verspreid voor in de stengel (zwarte pijlen in A). Elke vaatbundel is omgeven door een schede. Cambium betrokken in secundaire diktegroei ontbreekt hier (B). |
In tweezaadlobbigen zijn vaatbundels gerangschrikt als eilanden in een kring (oranje pijlen in C). Aanwezig is een cambiumlaag (stippenlijn in D) waaruit secundaire diktegroei zal ontstaan. |
 |  |
 |
 |
Maisplanten (helemaal links) en Palmbomen (hierlinks) zijn beide eenzaadlobbigen. In tegenstelling tot tweezaadlobbigen, zoals de zonnebloem hierrechts, kennen eenzaadlobbigen geen echte secundaire diktegroei. De toename in omvang van de stam bij de palmboom ontstaat vanuit een breed apicaal meristeem. |

|